


Savvas Christoforidis
Director of BRI
Group Leader (Researcher A’ level), Professor of Biological Chemistry
Our group studies the interplay between endocytosis, signalling and exocytosis in endothelial cells, focusing on the molecular mechanisms and their role in vascular physiology.
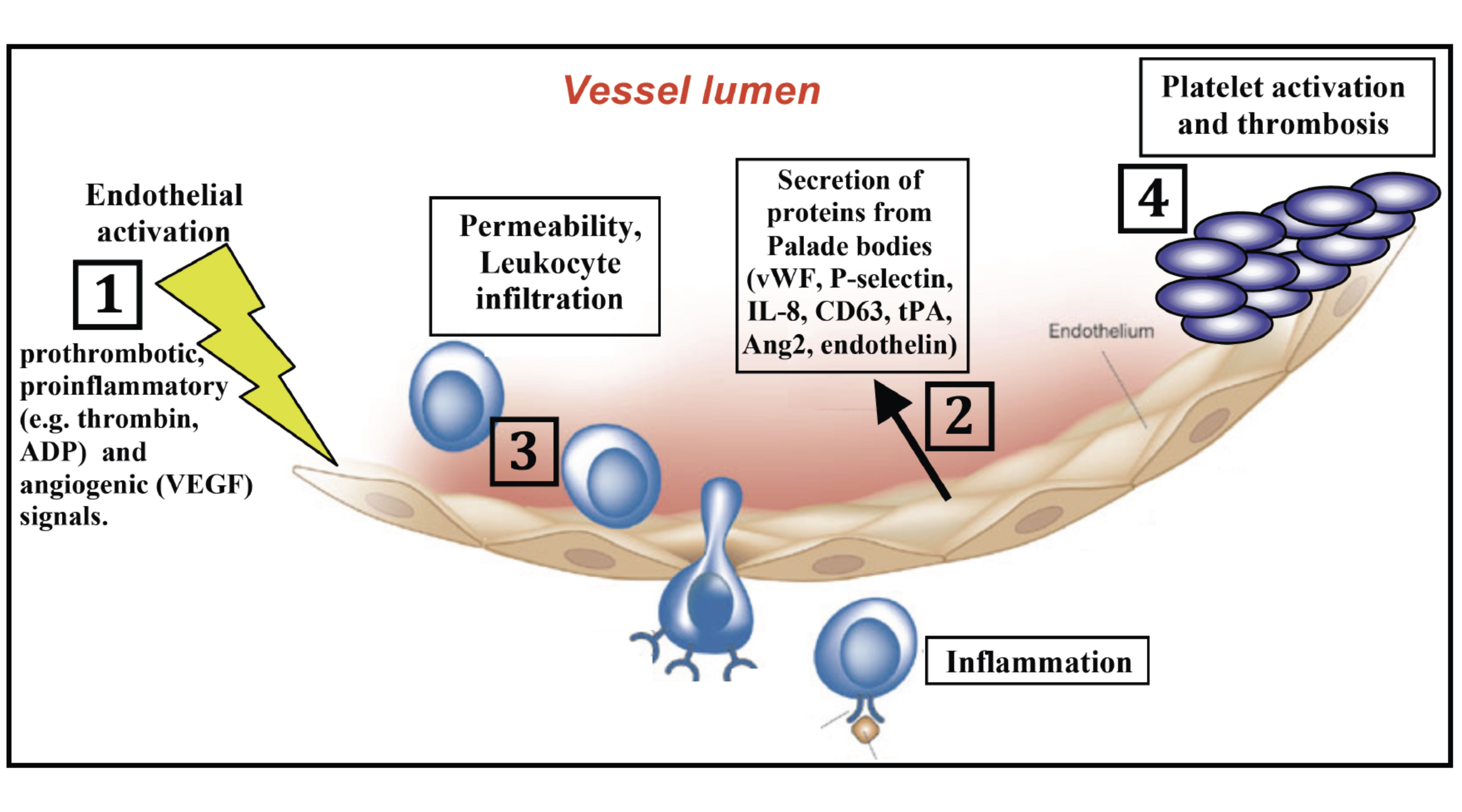
The inner wall of blood vessels is covered by endothelial cells, which play critical roles in the majority of fatal illnesses, including cardiovascular diseases, inflammatory disorders and neo-angiogenesis in cancer (see Fig 1. below). However, the exact mechanisms that govern, 1st, the properties of healthy endothelium, and 2nd, endothelial dysfunction in vascular diseases, are only poorly understood.
Intriguingly, key endothelial molecules that play important role in these pathophysiological processes are stored in specialized organelles called Weibel Palade bodies. Upon activation of endothelial cells, Weibel Palade bodies travel to the cell surface and fuse with plasma membrane, thereby releasing they cargo molecules in the blood stream. Among the secreted proteins are several key players in inflammation, thrombosis, angiogenesis and vessel tone, such as P-selectin, von willebrand factor (vWF), Angiopoeitin-2, endothelin, and others (see Fig.1 below).
One of the most important activators of Weibel Palade body secretion is VEGF (Vascular Endothelial Growth Factor), a growth factor that plays important role in tumour angiogenesis. We previously found that constitutive endocytosis protects the receptor of VEGF (VEGR2) against shedding (Basagiannis D and Christoforidis S, J Biol Chem, 2016, 291, 16892-16903), while addition of VEGF reroutes VEGFR2 towards macropinocytosis, a critical event for VEGF-induced signalling in vitro and for angiogenesis in vivo (Basagiannis D et al., J Cell Sci, 2016, 129, 4091-4104; Basagiannis D. et al., Sci. Rep., 2017, Mar 22;7:45035).
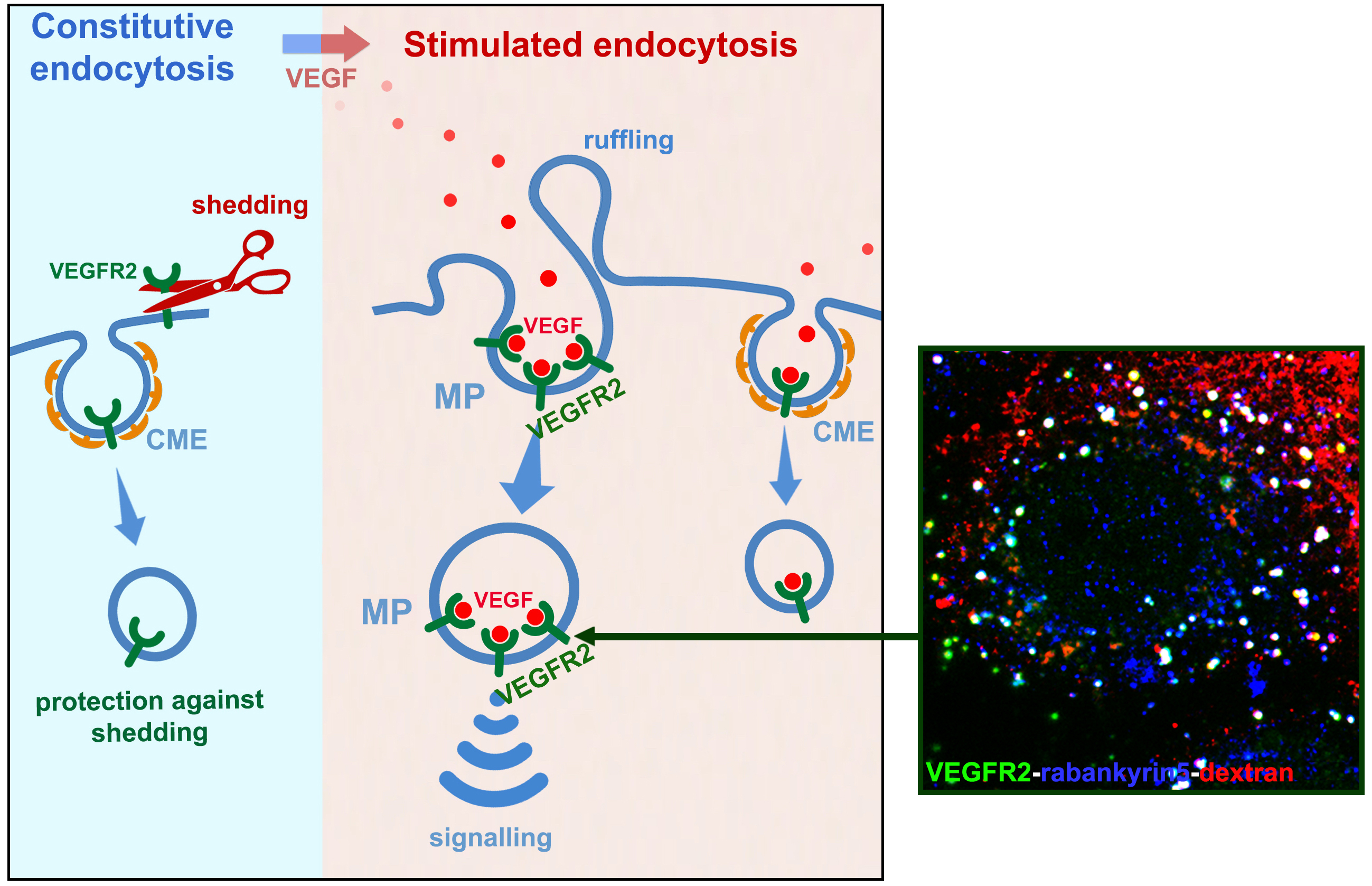
Following these findings, we are currently aiming in elucidating the mechanism by which macropinocytosis controls VEGFR2 signalling and how the signals are conveyed to Weibel Palade bodies, for triggering exocytosis (recently funded by an HFRI grant for Academic Staff and Researchers).
Overall, our group is interested in understanding the interconnections between endocytosis, signalling and exocytosis in endothelial cells and their importance in vascular physiology. Ultimately, the findings of our studies will contribute to the design of more effective therapeutic approaches in vascular diseases.
Role of endothelium in Alzheimer’s disease
In our effort to get deeper insights into the role of endothelium in vascular physiology, we have recently initiated a new project aiming to reveal the role of brain endothelium (the so-called blood brain barrier, BBB) in Alzheimer’s disease (recently funded by the Horizon Europe MSCA PF programme, to the Post-doctoral fellow Eftychia Vasili; “ClearPath” project). In this context, we focus to uncover how the brain’s blood vessels clear toxic amyloid-beta (Aβ) peptides, a key factor in the development of Alzheimer’s disease (AD). We have developed a novel human blood-brain barrier (BBB) transwell model from patient-derived stem cells, which allows us to study how different cellular uptake pathways contribute to Aβ clearance. By combining molecular and cellular biology, bioinformatics analysis of large patient datasets, and validation in animal models including C. elegans and mice, we aim to map the specific mechanisms by which Aβ is transported out of the brain. The ultimate goal is to identify new targets for therapies that prevent Aβ accumulation and slow the progression of Alzheimer’s disease.
Other projects in the lab include:
- Mechanism of overactivation of oncogenic PI3Kinase and screening for novel inhibitors (project in close collaboration with teams from BRFAA/Athens). Ref: Papafotika A, et al and Christoforidis S. “A new functional assay reveals that membrane binding is critical for overactivation of the phosphoinositide 3-kinase H1047R mutant.” J Biol Chem. 2026 doi: 10.1016/j.jbc.2026.111207
- The role of new interactors of the small GTPase Rab5 in membrane trafficking and signalling (in collaboration with the group of Marino Zerial, MPI-CBG/Dresden)
- Investigating the role of growth factor receptor internalization in the differentiation of human stem cells into vascular lineages
Methodology
To achieve the objectives of the projects described above, besides using standard techniques in the field of Biochemistry, Molecular and Cell Biology, we employ leading edge technology such as super resolution STED confocal microscopy, Total Internal Reflection Fluorescence Microscopy (TIRF-M), quantitative image analysis and proteomics by high resolution mass spectrometry.



